Boundary Activity
On this page the ‘emotion processing’ activity within individualized networks will be examined alongside the activity in the the amygdala, as well as in a set of meta-analytic ROIs, and group-based networks.
Primary Question: Do the individualized networks exhibit significant ‘emotion processing’ activity?
Reminder: We will only consider ‘emotion processing’ activity that is the result of more positive activity during the faces condition as compared to the shapes condition. Any activity that is the result of less negative activity during the faces condition as compared to the shapes condition will not be highlighted below.
Boundary Activity - Results:
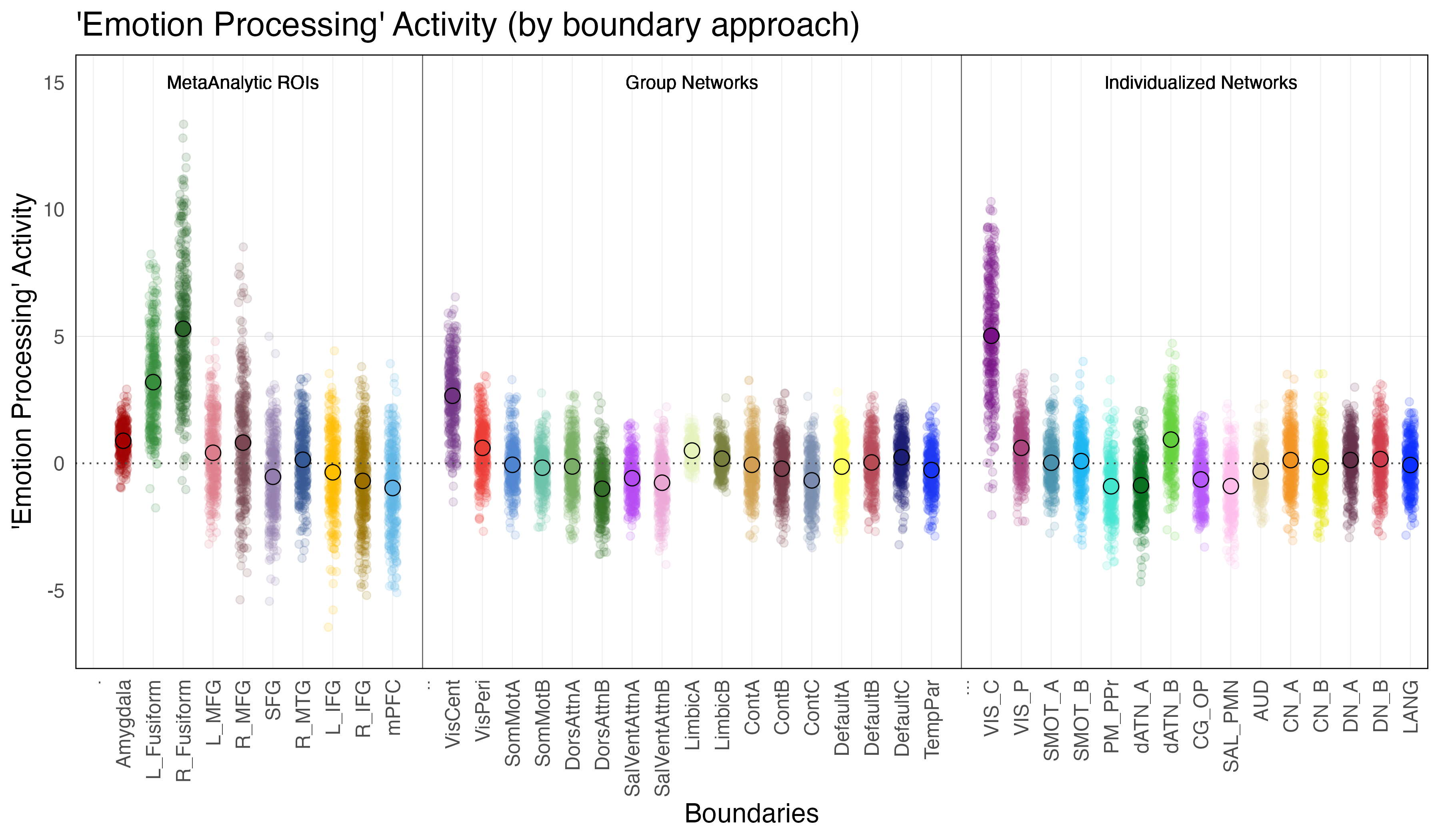
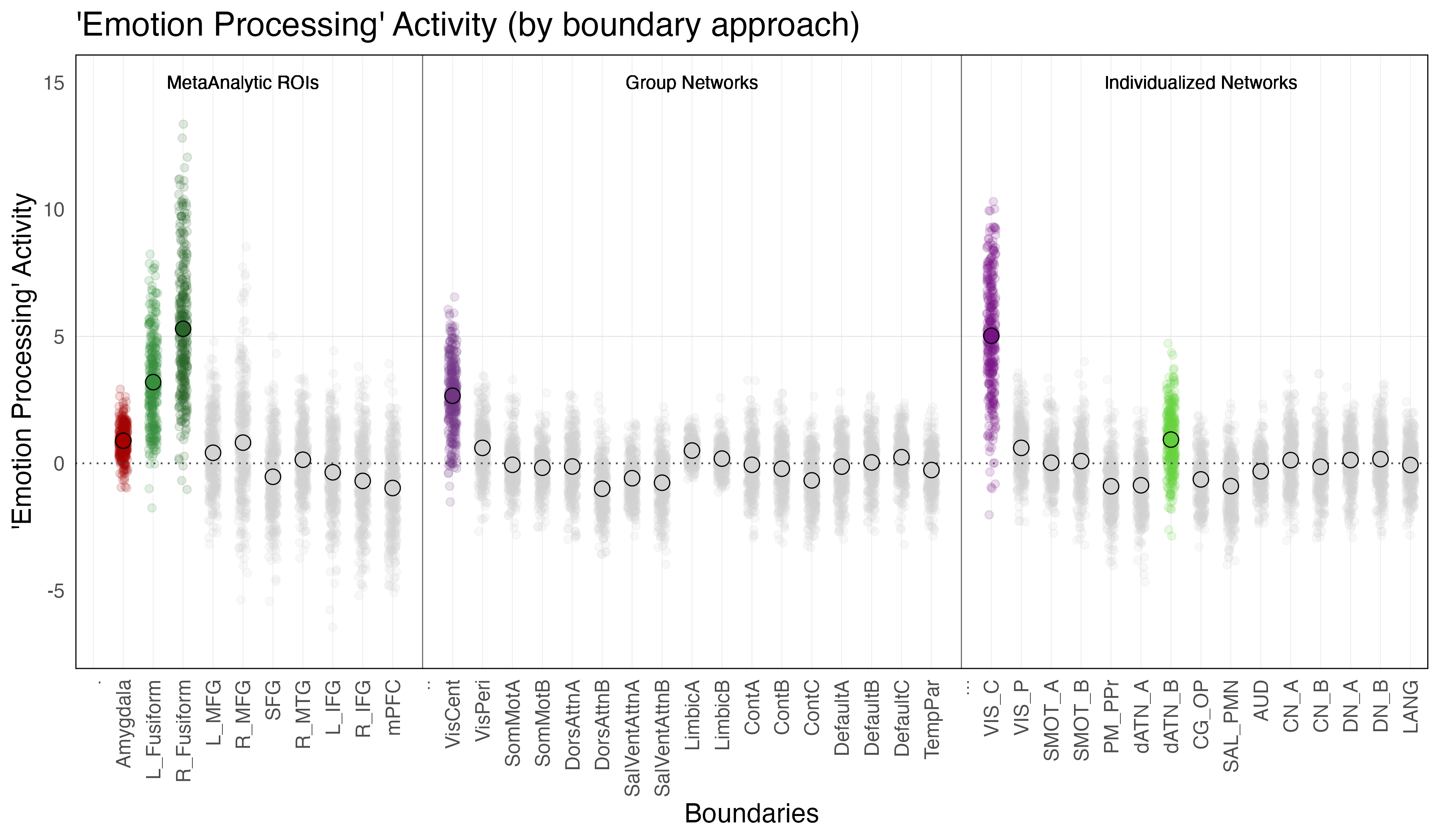
Amygdala. The amygdala exhibited significant ‘emotion processing’ activity (b = 0.89; SE = 0.12, p < 0.0001). Though the amygdala exhibited significant ‘emotion processing’ activity, examination of amygdala activity during the faces and shape conditions independently reveals that this ‘emotion processing’ activity is the result of only a slight increase in activity above zero during the faces condition and a slight decrease below zero during the shapes condition.
Meta-Analytic Regions of Interest (ROIs). Two meta-analytic regions of interest (ROIs) exhibited significant ‘emotion processing’ activity: left fusiform (b = 3.2, SE = 0.13, p < 0.0001) and right fusiform ((b = 5.3, SE = 0.13, p < 0.0001). It is interesting to note the laterilization in activity between the right and left fusiform, with the right fusiform exhibiting notably more activity that the left. (Note: left MFG, right MFG and right MTG technically exhibited significant ‘emotion processing’ activity, but this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations).
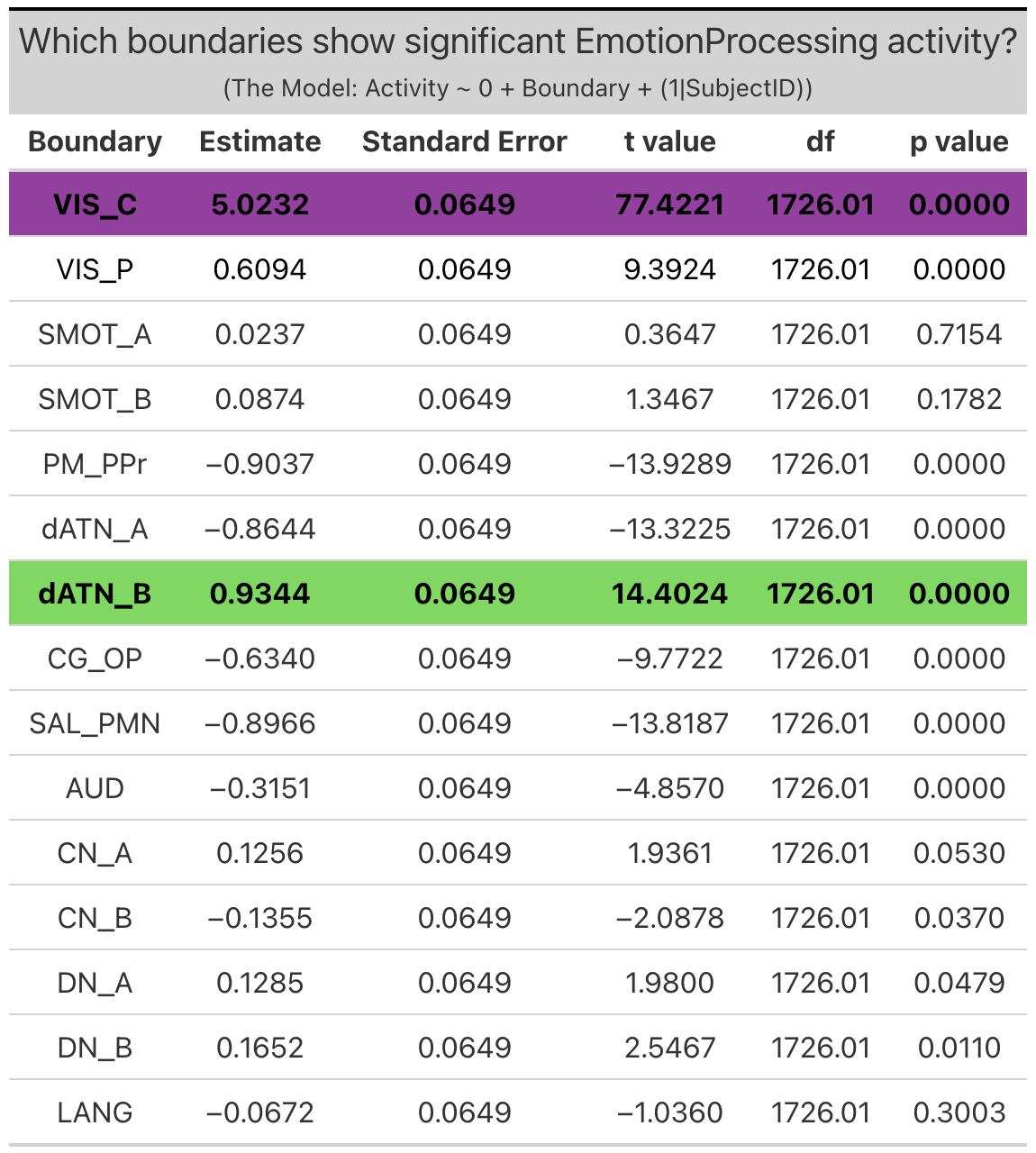
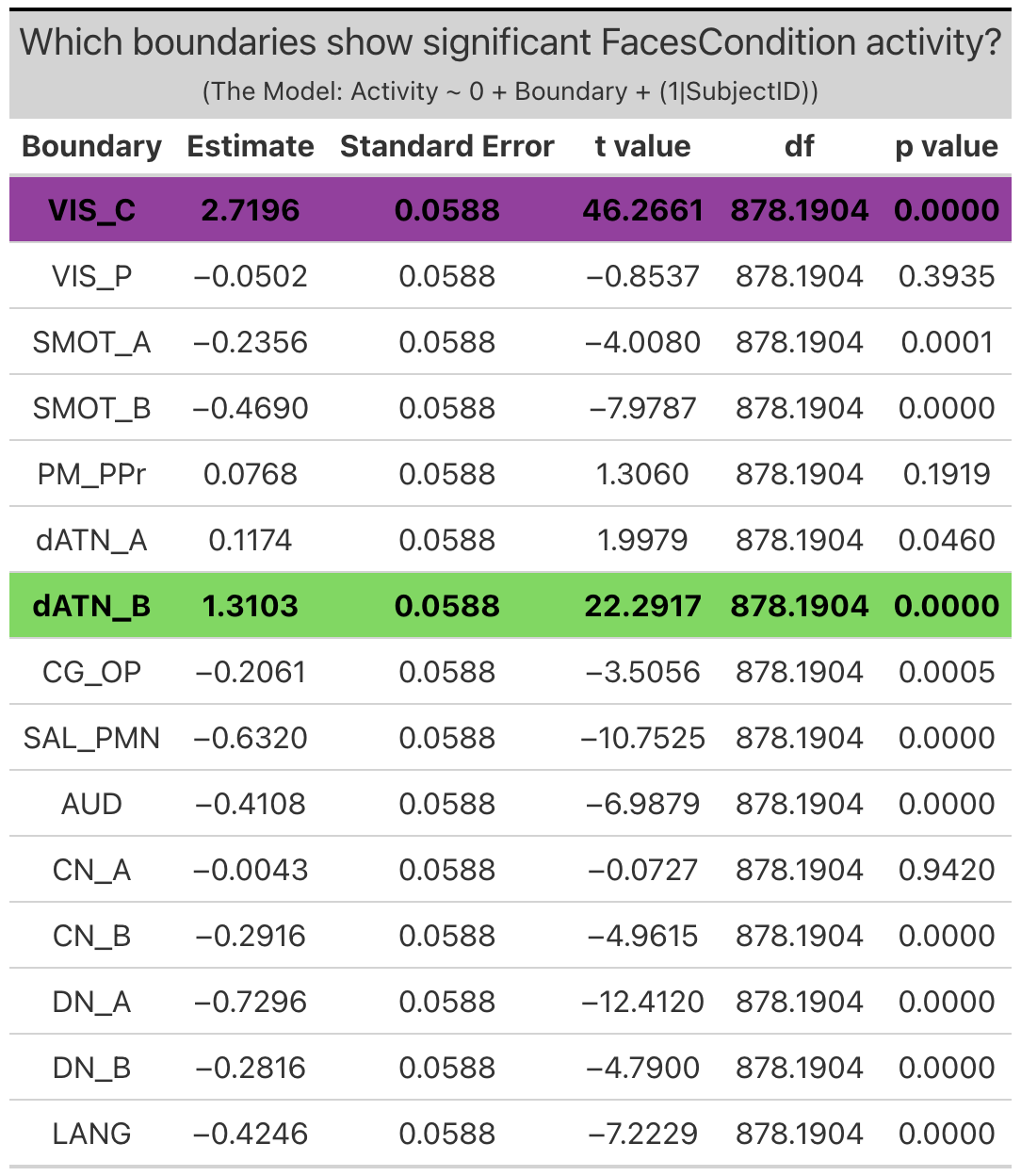
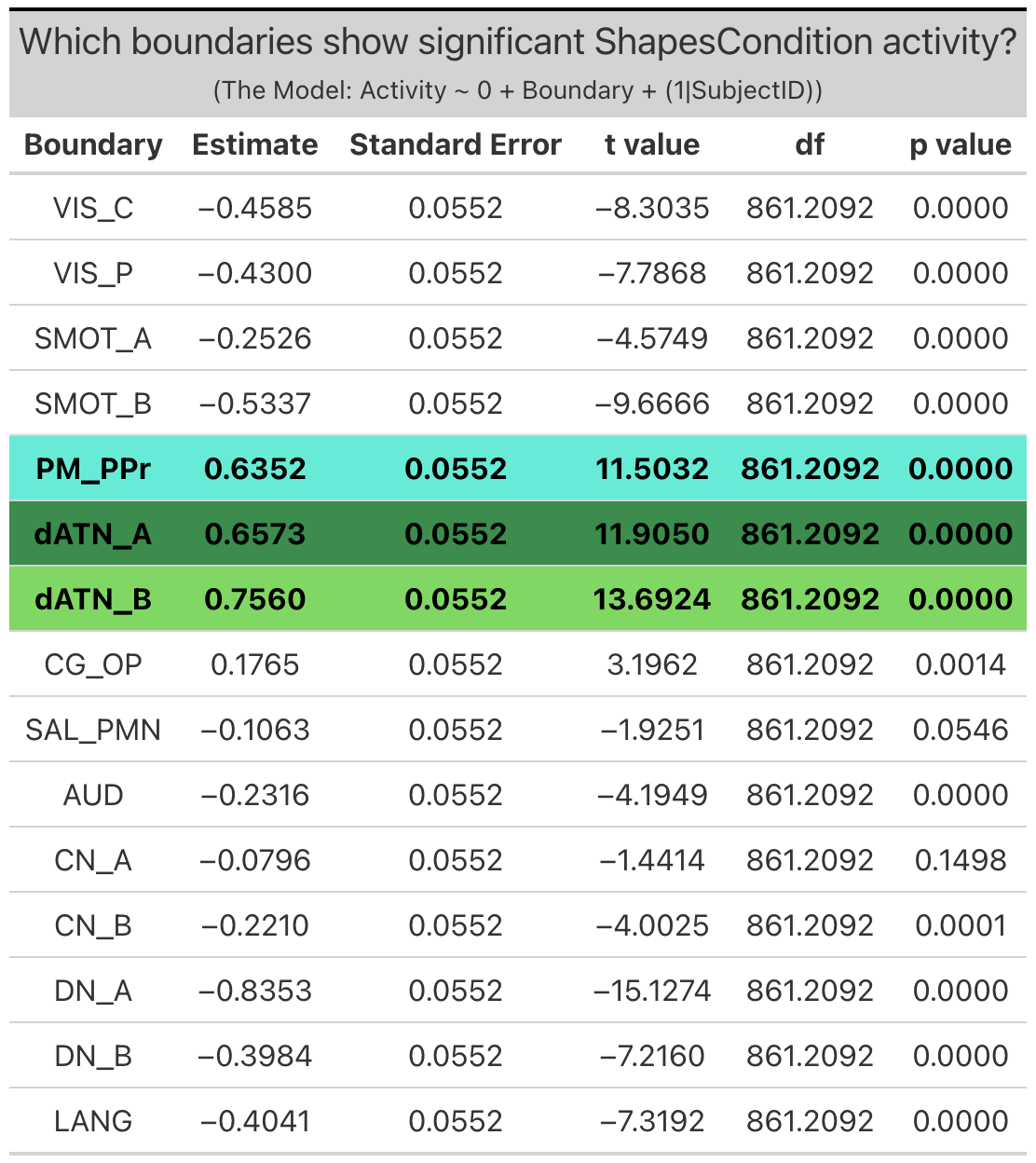
Individualized Networks. Two individualized networks exhibited significant ‘emotion processing’ activity: the visual-central network (VIS-C) (b = 5.0, SE = 0.065, p < 0.0001), and dorsal attention network B (dATN-B) (b = 0.93, SE = 0.065, p < 0.0001). (Note: visual-peripheral network (VIS-P), but again this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations).
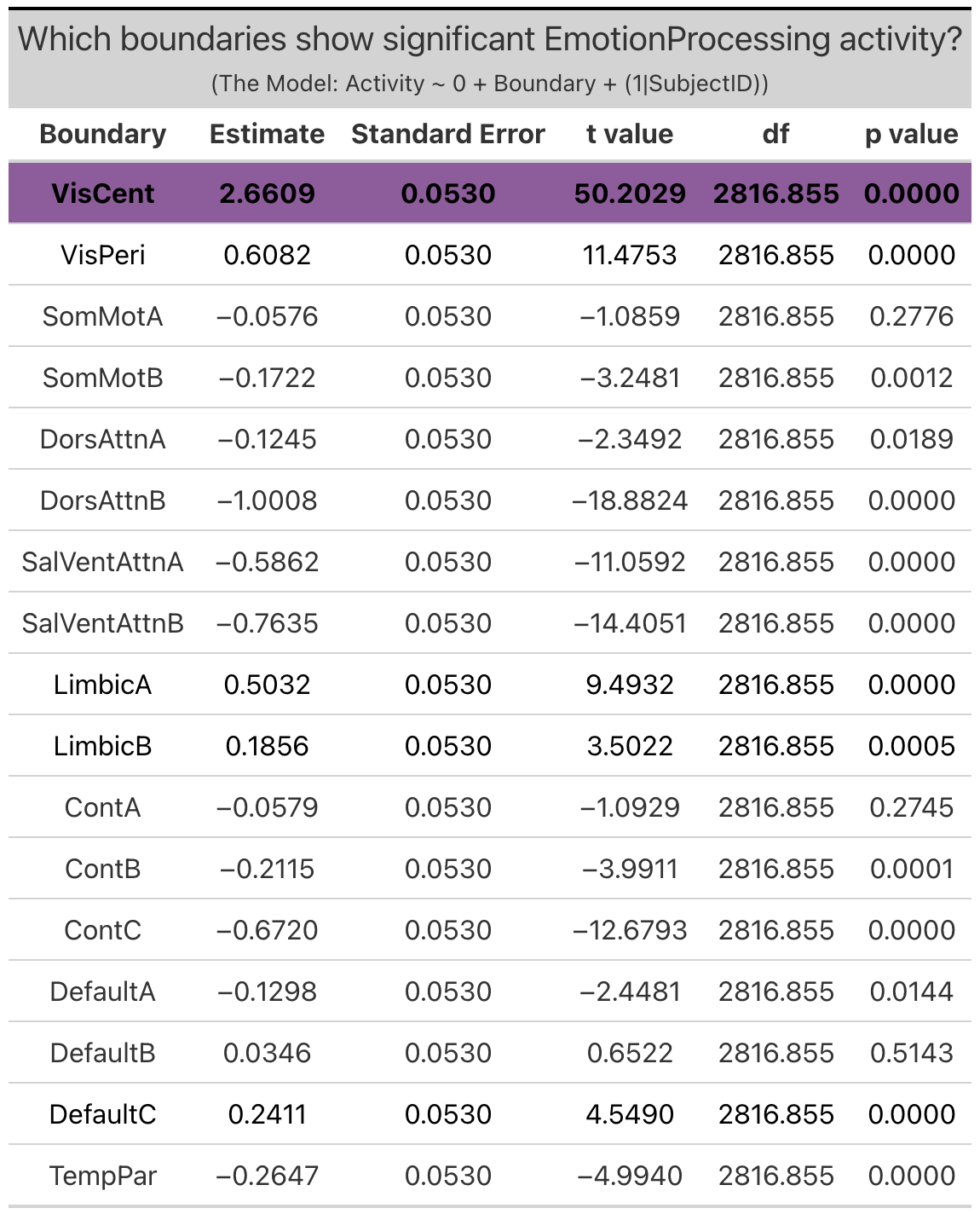
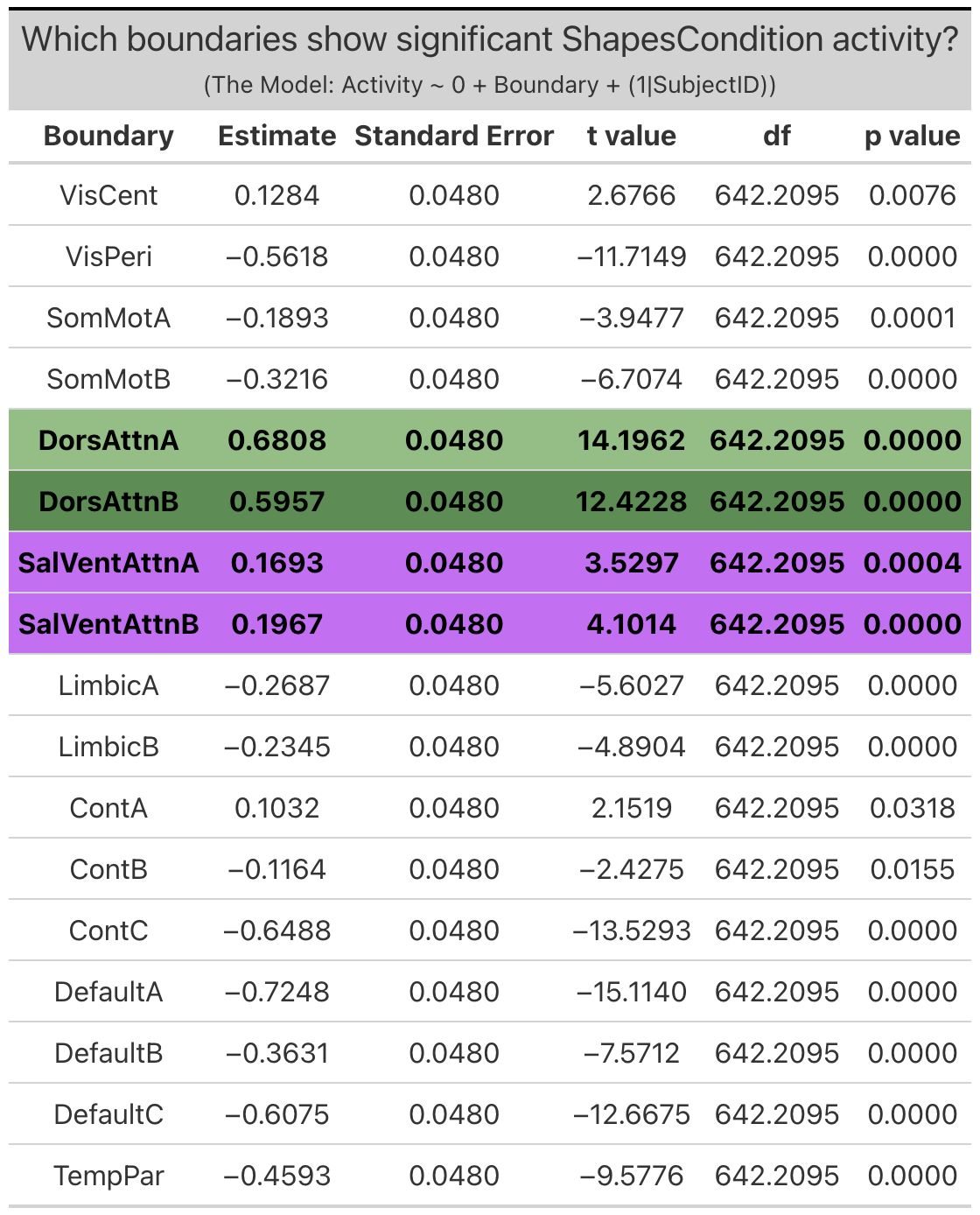
Group Networks. The only group network that exhibited significant ‘emotion processing’ activity was the visual-central network (VisCent) (b = 2.7, SE = 0.053, p < 0.0001). (Note: visual-peripheral network (VisPeri), default network C (DefaultC), limbic A and limbic B technically also exhibited significant ‘emotion processing’ activity, but again this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations). Given the effects observed within the dorsal attention network B of the individualized networks, we additionally reviewed activity within dorsal attention A of the group networks, the topological equilivent network despite alternate naming convention. Reviewing the face and shapes condition activity independently revealed significant activity within dorsal attention network a (DorsAttnA) for both the faces condition (b = 0.59, SE = 0.049, p < 0.0001) and the shapes condition (b = 0.68,SE = 0.048, p < 0.0001)). However, this activity was not meaningfully different between conditions, unlike in the individualized networks, where dorsal attention network B (dATN-B) exhibited significant ‘emotion processing’ activity given increased activation during the faces condition as compared to the shapes condition.
Results Tables
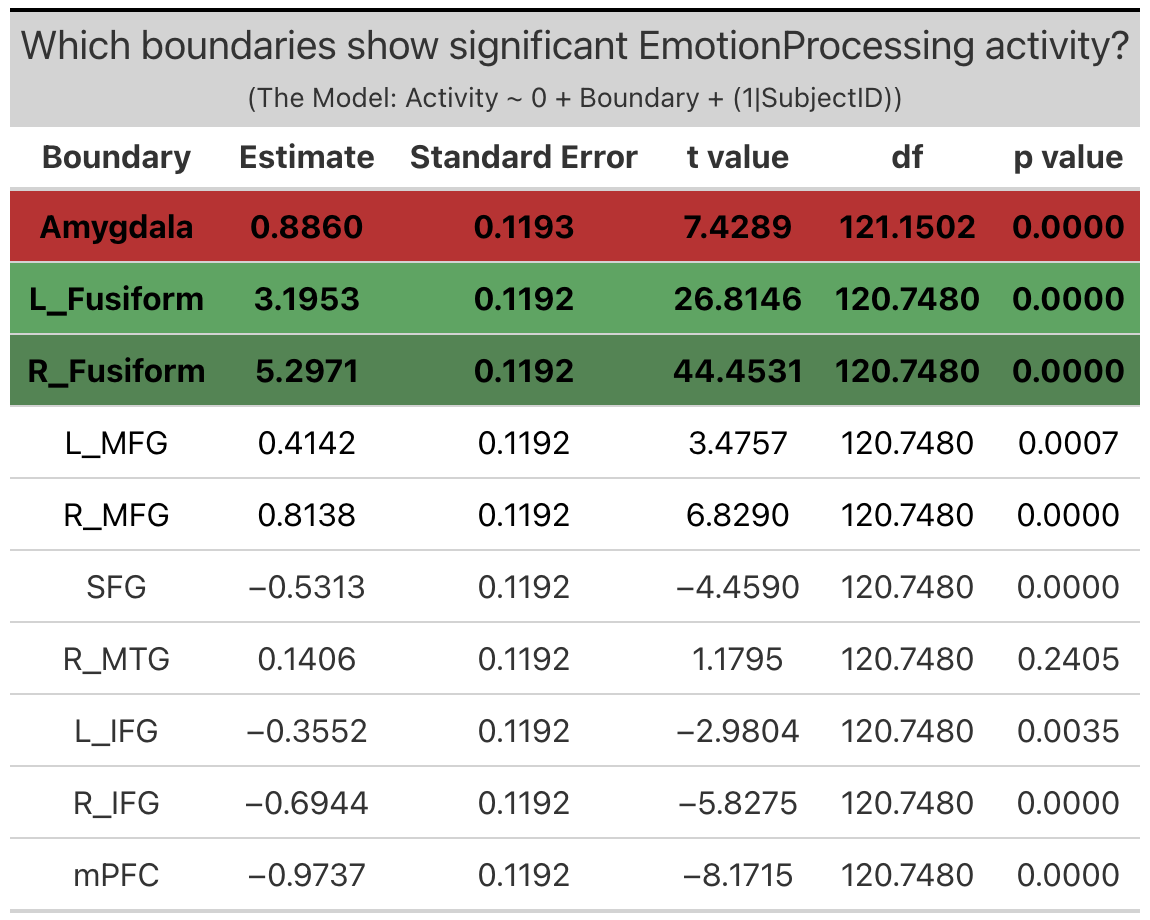
Amygdala. The amygdala exhibited significant ‘emotion processing’ activity (b = 0.89; SE = 0.12, p < 0.0001). Though the amygdala exhibited significant ‘emotion processing’ activity, examination of amygdala activity during the faces and shape conditions independently reveals that this ‘emotion processing’ activity is the result of only a slight increase in activity above zero during the faces condition and a slight decrease below zero during the shapes condition.
Meta-Analytic Regions of Interest (ROIs). Two meta-analytic regions of interest (ROIs) exhibited significant ‘emotion processing’ activity: left fusiform (b = 3.2, SE = 0.13, p < 0.0001) and right fusiform ((b = 5.3, SE = 0.13, p < 0.0001). It is interesting to note the laterilization in activity between the right and left fusiform, with the right fusiform exhibiting notably more activity that the left. (Note: left MFG, right MFG and right MTG technically exhibited significant ‘emotion processing’ activity, but this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations).
Individualized Networks. Two individualized networks exhibited significant ‘emotion processing’ activity: the visual-central network (VIS-C) (b = 5.0, SE = 0.065, p < 0.0001), and dorsal attention network B (dATN-B) (b = 0.93, SE = 0.065, p < 0.0001). (Note: visual-peripheral network (VIS-P), but again this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations).
Group Networks. The only group network that exhibited significant ‘emotion processing’ activity was the visual-central network (VisCent) (b = 2.7, SE = 0.053, p < 0.0001). (Note: visual-peripheral network (VisPeri), default network C (DefaultC), limbic A and limbic B technically also exhibited significant ‘emotion processing’ activity, but again this activity was the result of less negative activity during the faces condition as compared to the shapes condition, and so will not be considered in further investigations). Given the effects observed within the dorsal attention network B of the individualized networks, we additionally reviewed activity within dorsal attention A of the group networks, the topological equilivent network despite alternate naming convention. Reviewing the face and shapes condition activity independently revealed significant activity within dorsal attention network a (DorsAttnA) for both the faces condition (b = 0.59, SE = 0.049, p < 0.0001) and the shapes condition (b = 0.68,SE = 0.048, p < 0.0001)). However, this activity was not meaningfully different between conditions, unlike in the individualized networks, where dorsal attention network B (dATN-B) exhibited significant ‘emotion processing’ activity given increased activation during the faces condition as compared to the shapes condition.
Results Tables
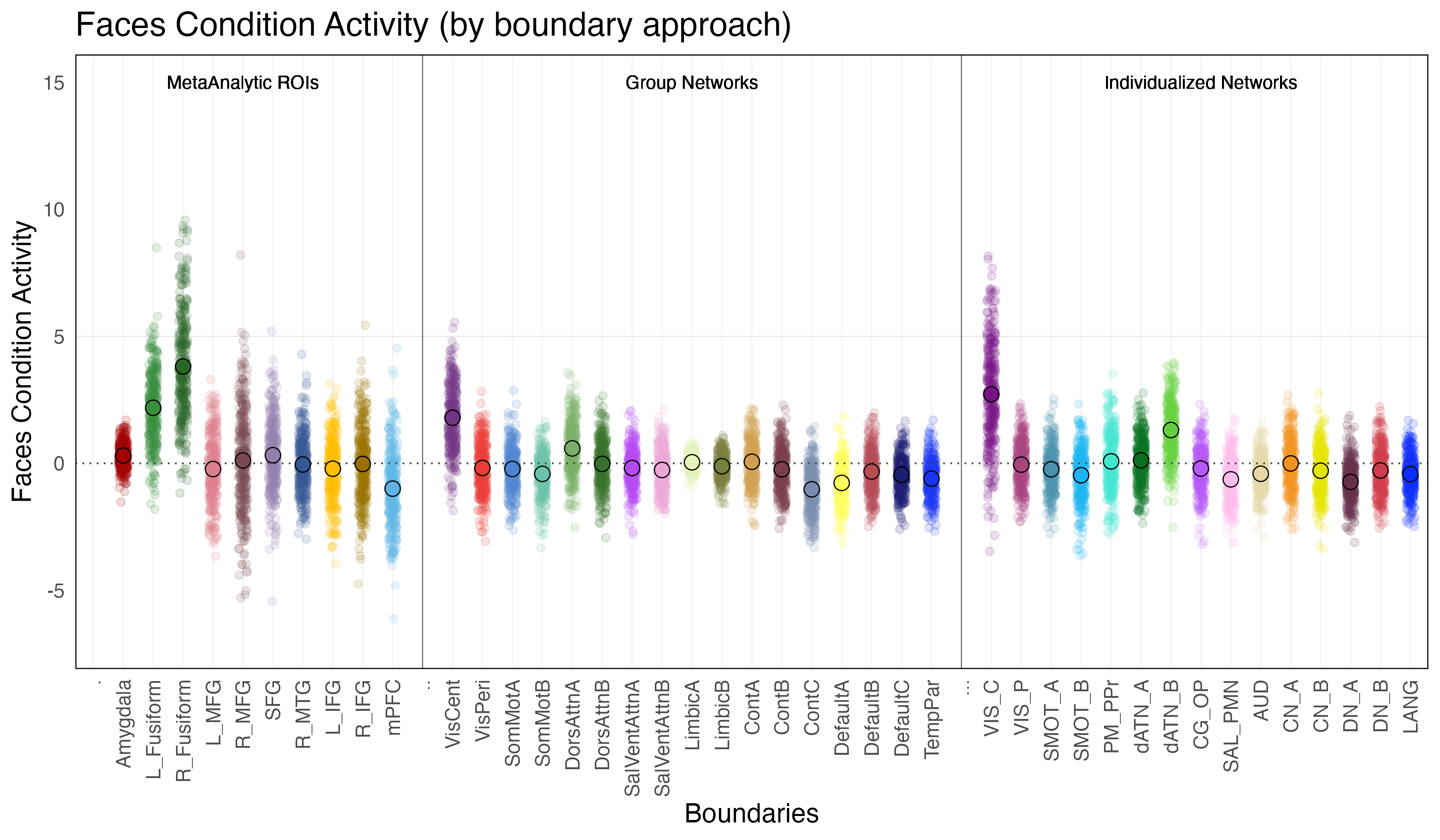
Faces Condition Activity is defined as the difference in activation during the faces condition relative to ‘baseline’ (ie. when the subjects are observing a fixation cross in between trials).
Results Tables
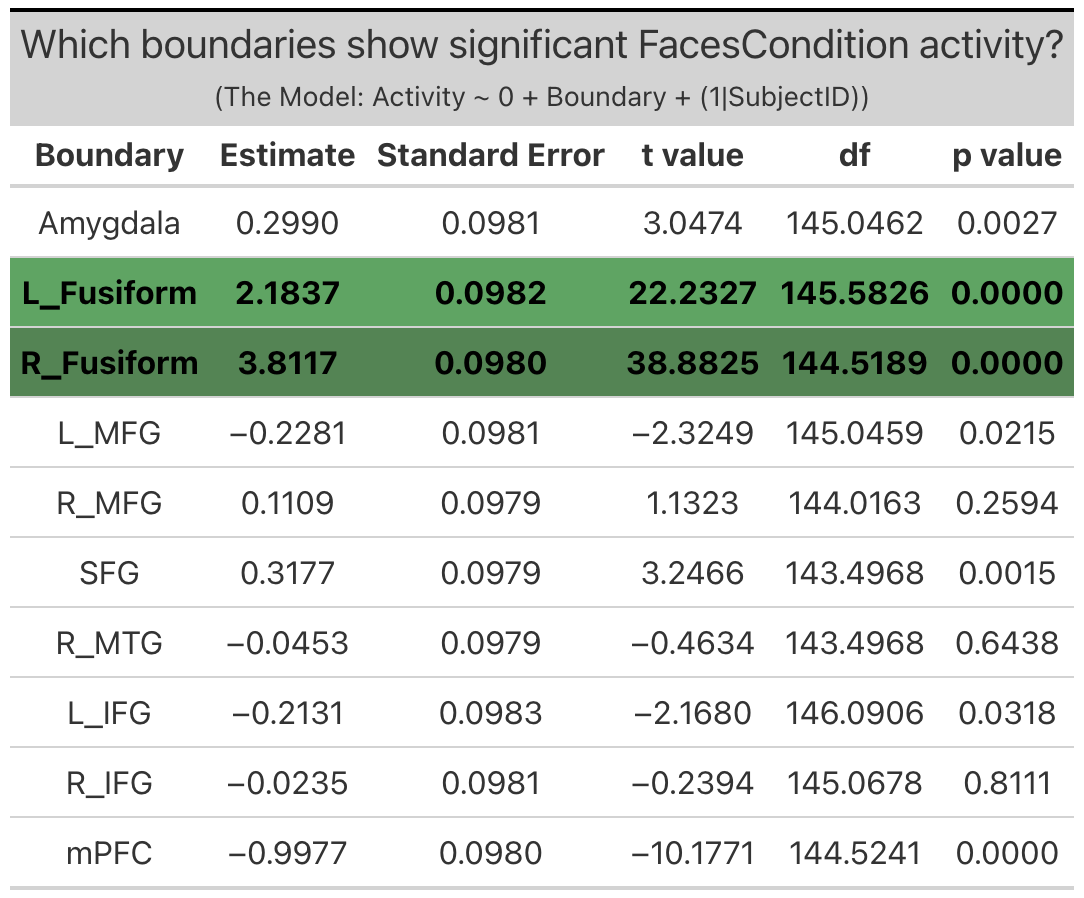
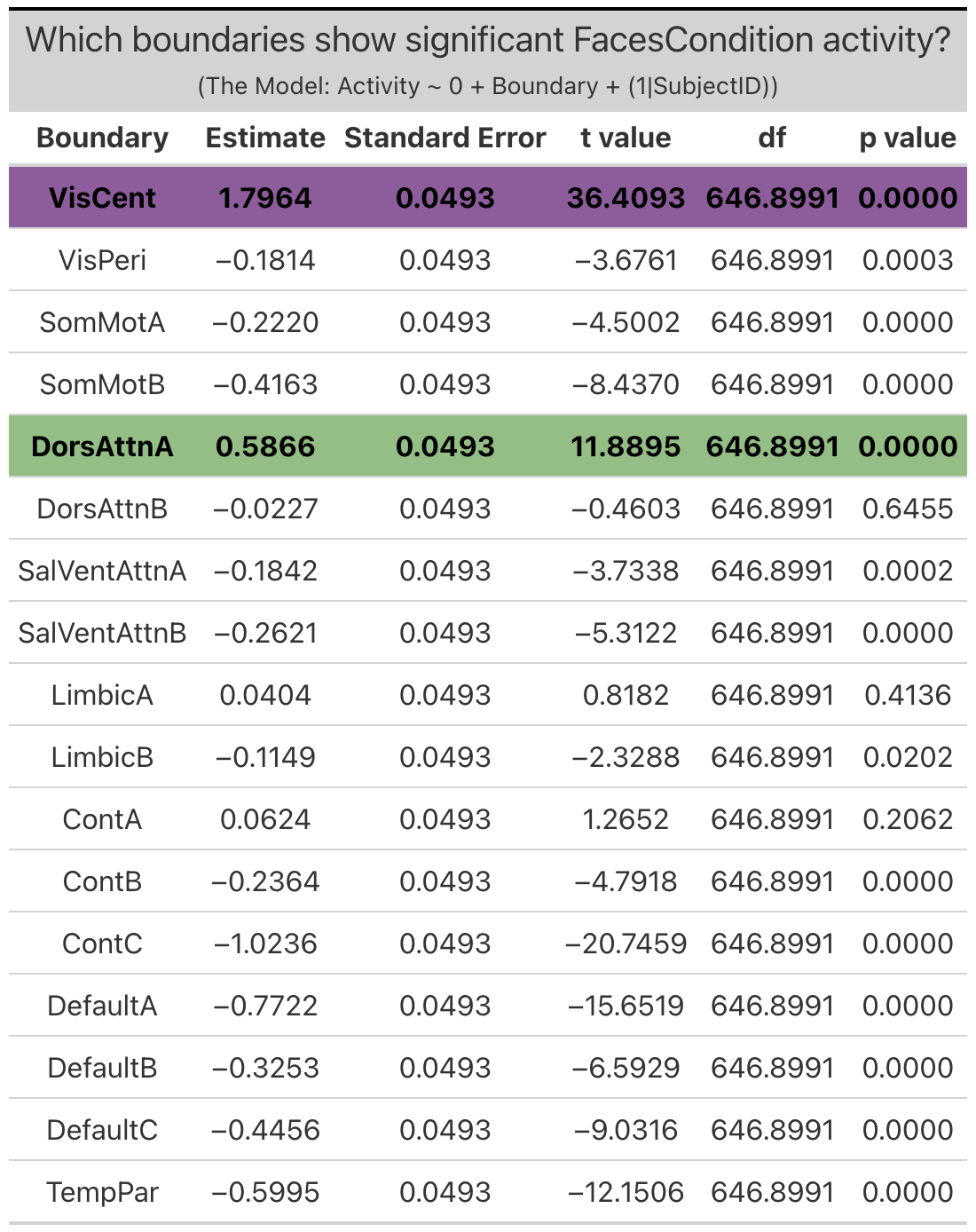
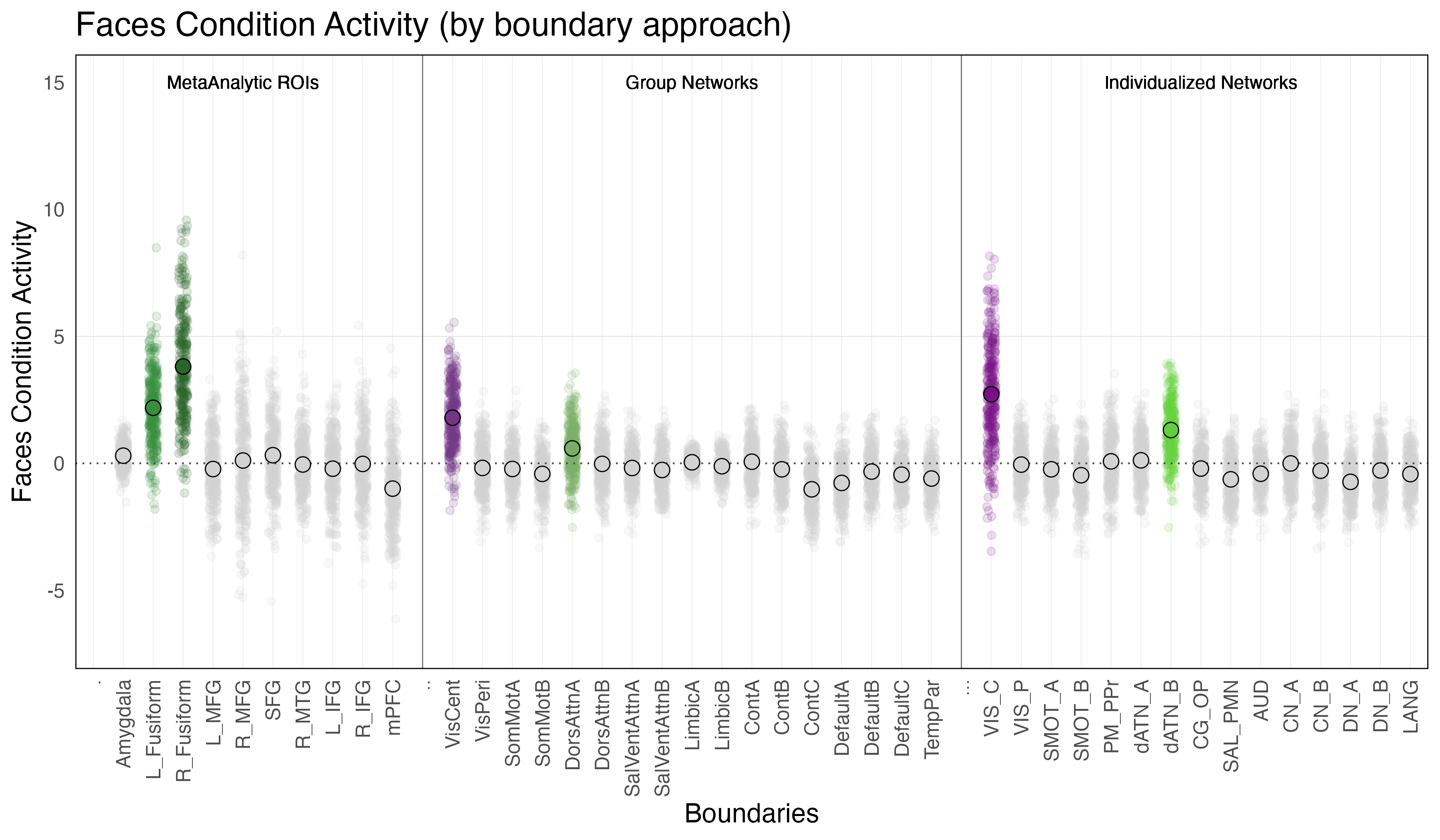
Faces Condition Activity is defined as the difference in activation during the faces condition relative to ‘baseline’ (ie. when the subjects are observing a fixation cross in between trials).
Results Tables
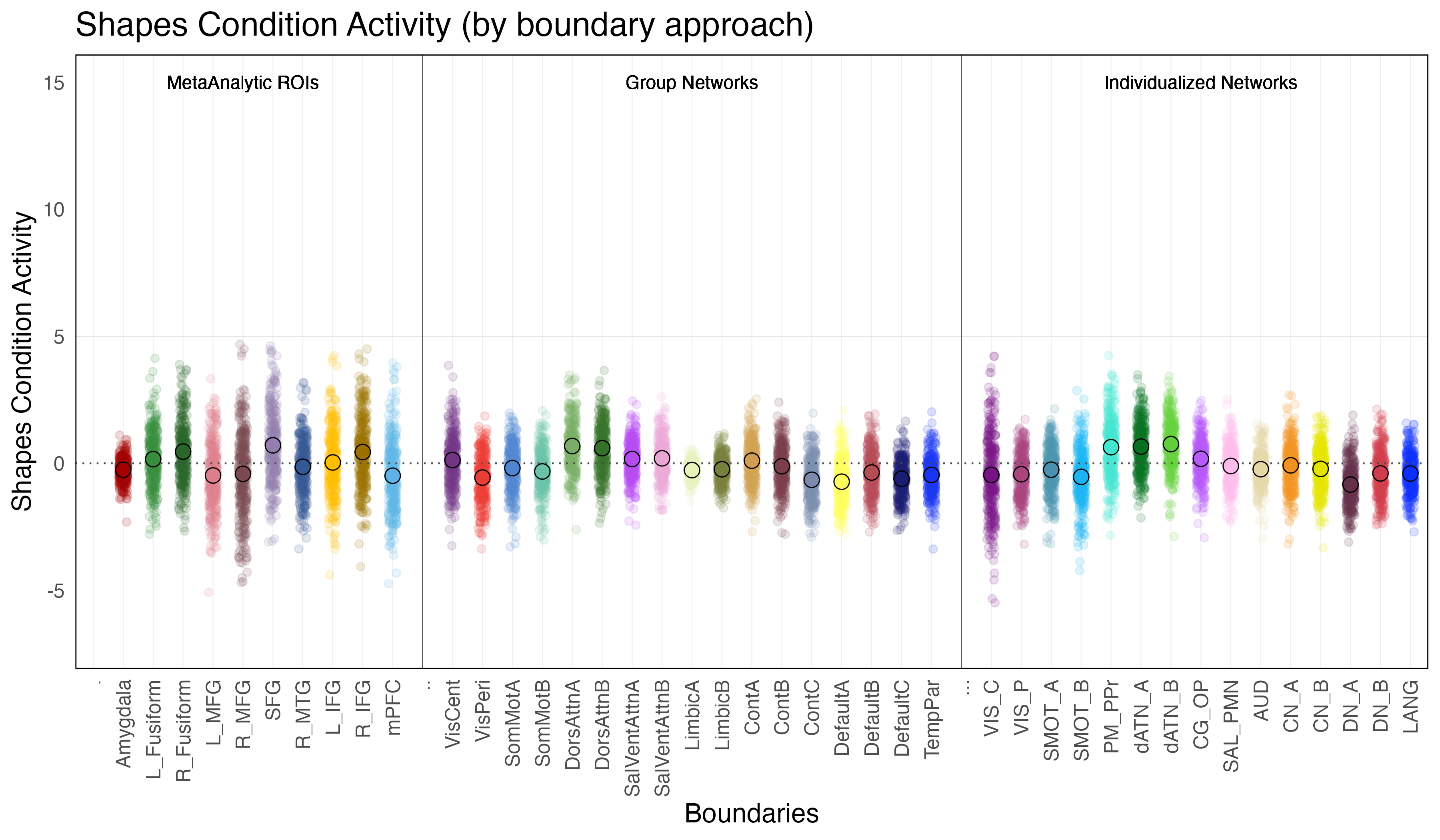
Shapes Condition Activity is defined as the difference in activation during the shapes condition relative to baseline (ie. when the subjects are observing a fixation cross in between trials).
Results Tables
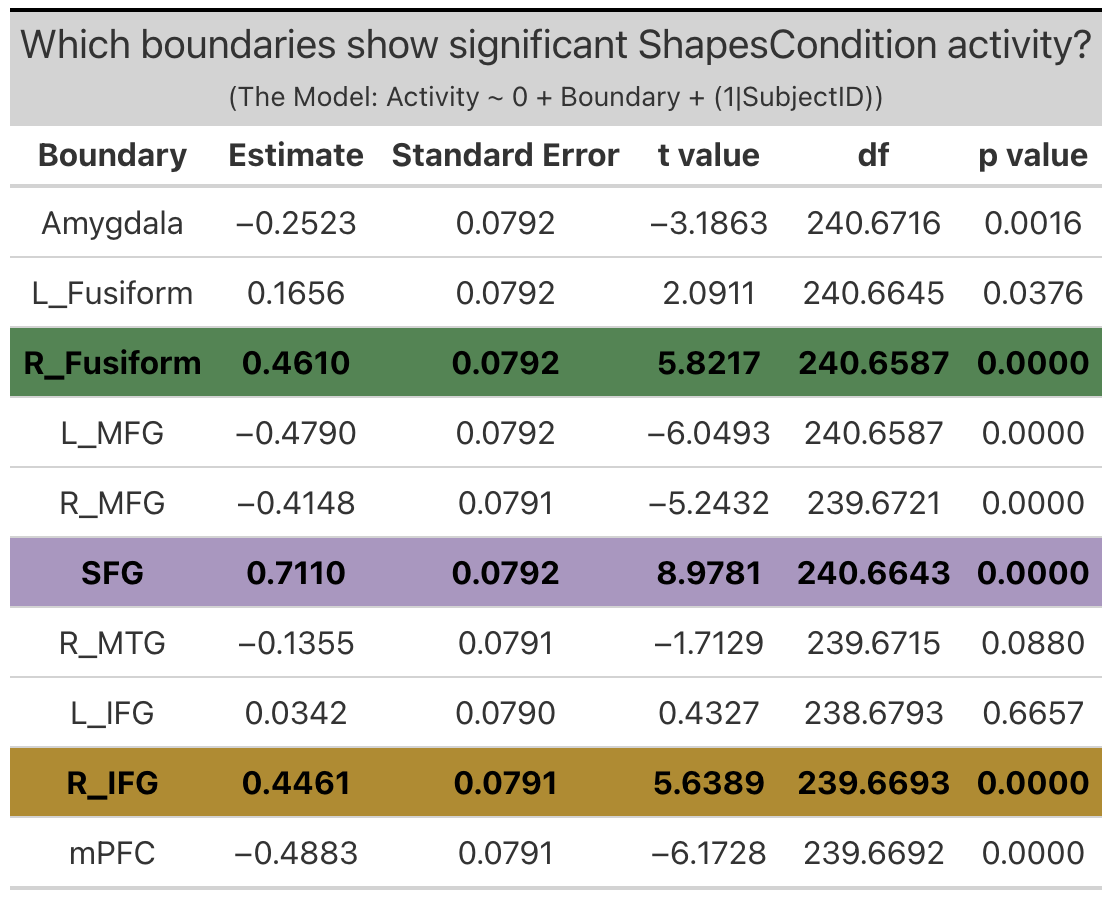
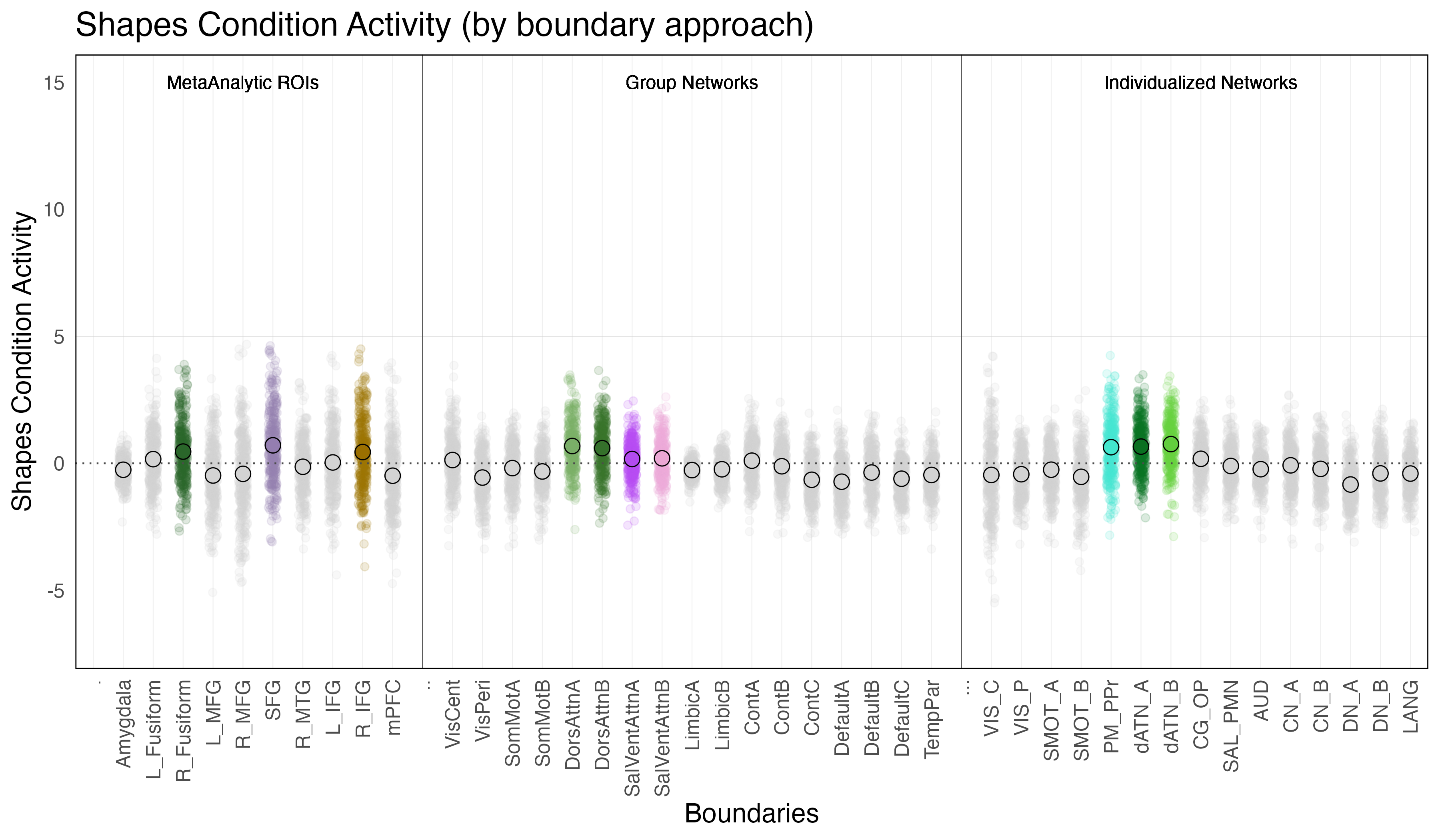
Shapes Condition Activity is defined as the difference in activation during the shapes condition relative to baseline (ie. when the subjects are observing a fixation cross in between trials).
Results Tables
Summary of Observations — Boundary Activity
In sum, significant ‘emotion processing’ activity was observed at both a regional level (amygdala, left and right fusiform), as well as at a network-level (visual-central network and dorsal attention network B (of the individualized networks)). The fusifrom and the individualized visual-central network showed comparable levels of ‘emotion processing’ activity. Significant activity was also observed within the visual-central network of the group-based parcellation, however this activity was notably less than that observed in the individualized networks.
The fusiform falls within the ventral portion of the cortex that also comprises portions of both the visual-central network (VIS-C) and portions of the dorsal attention network B (dATN-B). It is therefore possible that the network activity observed simply reflects fusiform activity driving up the average network activity. To examine this possibility, the topological overlap between the left and right fusiform and the networks will be examined to confirm ROI placement relative to the networks. Following the topological comparison, activity of each vertex within each network will be examined to determine if the observed increase in network activity is the result of global increased in activity across all vertices in a network or the result of increases within only a subset of vertices (ie. a region that falls within a network, such as the fusiform falling with the visual-central network).
UP NEXT: Topological Comparison – Fusiform vs. Networks
Primary Question: Where does the fusiform fall within the network parcellations?